L’insuline, hormone produite par le pancréas, joue un rôle crucial dans la régulation du métabolisme. Cette protéine complexe orchestre une série de processus biochimiques essentiels à l’homéostasie énergétique de l’organisme. Son action principale consiste à faciliter l’absorption du glucose par les cellules, mais son influence s’étend bien au-delà, affectant le métabolisme des lipides et des protéines. Comprendre le fonctionnement de l’insuline est fondamental pour appréhender les mécanismes sous-jacents de nombreuses pathologies métaboliques, notamment le diabète.
Mécanisme d’action moléculaire de l’insuline
Structure et synthèse de l’insuline dans les cellules bêta pancréatiques
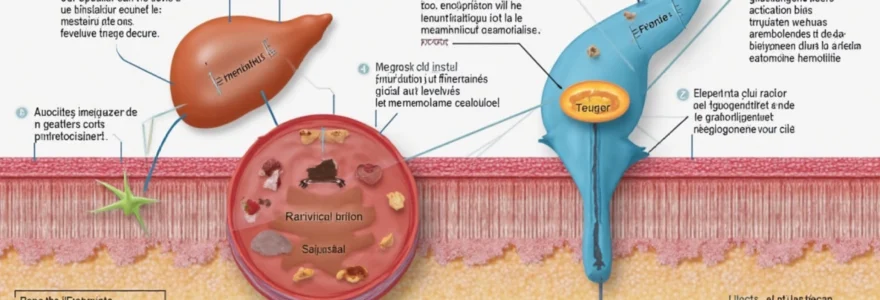
L’insuline est synthétisée au sein des cellules bêta des îlots de Langerhans du pancréas. Cette hormone peptidique se compose de deux chaînes polypeptidiques, A et B, reliées par des ponts disulfures. La biosynthèse de l’insuline débute par la production d’un précurseur appelé préproinsuline, qui subit ensuite des modifications post-traductionnelles pour former la proinsuline. Enfin, le clivage de la proinsuline donne naissance à l’insuline mature et au peptide C.
Le processus de sécrétion de l’insuline est finement régulé par la glycémie. Lorsque le taux de glucose sanguin augmente, les cellules bêta détectent ce changement et initient la libération d’insuline par exocytose. Cette réponse rapide permet un contrôle précis de la glycémie, essentiel au maintien de l’équilibre métabolique.
Liaison de l’insuline aux récepteurs membranaires INSR
Une fois libérée dans la circulation sanguine, l’insuline agit sur ses cellules cibles en se liant à des récepteurs spécifiques appelés INSR (Insulin Receptor). Ces récepteurs sont des protéines transmembranaires composées de deux sous-unités alpha extracellulaires et deux sous-unités bêta intracellulaires. La liaison de l’insuline aux sous-unités alpha déclenche un changement conformationnel qui active les sous-unités bêta, dotées d’une activité tyrosine kinase intrinsèque.
Cette activation des récepteurs INSR marque le début d’une cascade de signalisation intracellulaire complexe. La spécificité et l’affinité de la liaison insuline-récepteur sont cruciales pour la transmission précise du signal hormonal et l’initiation des effets métaboliques de l’insuline.
Cascade de signalisation intracellulaire via IRS et PI3K
L’activation des récepteurs INSR déclenche une série d’événements moléculaires en chaîne. Les substrats du récepteur de l’insuline (IRS) sont phosphorylés, créant des sites d’ancrage pour d’autres protéines de signalisation. Parmi celles-ci, la phosphatidylinositol 3-kinase (PI3K) joue un rôle central. Son activation conduit à la production de seconds messagers lipidiques, notamment le PIP3 (phosphatidylinositol-3,4,5-trisphosphate).
Cette cascade de signalisation implique de nombreuses kinases, dont la PDK1 et l’Akt/PKB. L’Akt, en particulier, est un régulateur clé des effets métaboliques de l’insuline, orchestrant diverses réponses cellulaires telles que la translocation des transporteurs de glucose et la régulation de la synthèse du glycogène.
Translocation des transporteurs GLUT4 vers la membrane cellulaire
L’un des effets les plus importants de la signalisation insulinique est la translocation des transporteurs de glucose GLUT4 vers la membrane plasmique. Dans les cellules musculaires et adipeuses, les GLUT4 sont normalement séquestrés dans des vésicules intracellulaires. Sous l’influence de l’insuline, ces vésicules fusionnent avec la membrane, augmentant ainsi la densité de GLUT4 à la surface cellulaire.
Ce processus de translocation des GLUT4 est essentiel pour l’absorption efficace du glucose sanguin par les tissus insulino-sensibles. Il représente le mécanisme principal par lequel l’insuline régule la glycémie, permettant une clairance rapide du glucose après un repas.
Régulation du métabolisme glucidique par l’insuline
Stimulation de l’absorption du glucose sanguin
L’effet le plus connu de l’insuline est sa capacité à stimuler l’absorption du glucose par les cellules. Cette action est particulièrement prononcée dans les tissus musculaires et adipeux, où l’insuline provoque une augmentation rapide et significative de l’entrée de glucose. Ce processus est médié principalement par la translocation des transporteurs GLUT4, qui agissent comme des portes d’entrée pour le glucose.
Dans le foie, l’insuline favorise également l’absorption du glucose, mais par un mécanisme différent. Les hépatocytes expriment principalement le transporteur GLUT2, qui n’est pas régulé par l’insuline de la même manière que GLUT4. Néanmoins, l’insuline augmente l’activité de la glucokinase hépatique, enzyme qui phosphoryle le glucose entrant, facilitant ainsi son absorption et son métabolisme.
Activation de la glycogénogenèse hépatique et musculaire
Une fois le glucose absorbé par les cellules, l’insuline oriente son devenir métabolique. Dans le foie et les muscles, elle stimule la glycogénogenèse, processus par lequel le glucose est converti en glycogène pour le stockage. Cette action implique l’activation de la glycogène synthase, enzyme clé de la synthèse du glycogène.
L’insuline active la glycogène synthase par deux mécanismes principaux : la déphosphorylation directe de l’enzyme et l’inhibition de la glycogène synthase kinase 3 (GSK3). Ces effets combinés augmentent considérablement la capacité des cellules à stocker le glucose sous forme de glycogène, créant ainsi une réserve énergétique importante.
Inhibition de la glycogénolyse et de la néoglucogenèse
Parallèlement à la stimulation du stockage du glucose, l’insuline inhibe les processus qui libèrent du glucose dans la circulation sanguine. Dans le foie, elle supprime la glycogénolyse (dégradation du glycogène) et la néoglucogenèse (production de glucose à partir de précurseurs non glucidiques).
L’inhibition de la glycogénolyse se fait principalement par l’inactivation de la glycogène phosphorylase, enzyme responsable de la dégradation du glycogène. Quant à la néoglucogenèse, l’insuline réduit l’expression des enzymes clés de ce processus, notamment la phosphoénolpyruvate carboxykinase (PEPCK) et la glucose-6-phosphatase. Ces actions coordonnées de l’insuline contribuent à maintenir la glycémie dans des limites physiologiques étroites.
Effets de l’insuline sur le métabolisme lipidique
Stimulation de la lipogenèse dans le tissu adipeux
L’action de l’insuline ne se limite pas au métabolisme glucidique ; elle exerce également une influence majeure sur le métabolisme lipidique. Dans le tissu adipeux, l’insuline stimule la lipogenèse, processus de synthèse des triglycérides à partir du glucose. Cette action implique l’activation de plusieurs enzymes lipogéniques, notamment l’acétyl-CoA carboxylase et la fatty acid synthase.
De plus, l’insuline augmente l’expression et l’activité de la lipoprotéine lipase (LPL) à la surface des adipocytes. La LPL hydrolyse les triglycérides circulants dans les lipoprotéines, permettant ainsi l’absorption des acides gras libérés par les cellules adipeuses. Ce mécanisme contribue à l’accumulation de lipides dans le tissu adipeux après un repas.
Inhibition de la lipolyse et de la cétogenèse
Parallèlement à son action lipogénique, l’insuline exerce un puissant effet anti-lipolytique. Elle inhibe la lipase hormono-sensible, enzyme clé de la dégradation des triglycérides stockés dans les adipocytes. Cette inhibition réduit la libération d’acides gras libres dans la circulation, limitant ainsi leur utilisation comme source d’énergie par d’autres tissus.
Dans le foie, l’insuline supprime la cétogenèse, processus de production de corps cétoniques à partir des acides gras. Cette action est particulièrement importante en période postprandiale, lorsque l’organisme dispose d’un apport énergétique suffisant sous forme de glucose. L’inhibition de la cétogenèse contribue à prévenir l’accumulation excessive de corps cétoniques, qui peuvent être dangereux à haute concentration.
Régulation du métabolisme du cholestérol
L’insuline joue également un rôle dans la régulation du métabolisme du cholestérol. Elle stimule l’activité de l’enzyme HMG-CoA réductase, étape limitante de la biosynthèse du cholestérol. Cependant, l’insuline augmente aussi l’expression des récepteurs aux LDL (lipoprotéines de basse densité) dans le foie, favorisant ainsi la clairance du cholestérol sanguin.
Ces effets complexes de l’insuline sur le métabolisme lipidique illustrent son rôle central dans la coordination du stockage et de l’utilisation des nutriments. Un déséquilibre dans ces actions peut contribuer au développement de troubles métaboliques tels que la dyslipidémie et l’athérosclérose.
Rôle de l’insuline dans l’anabolisme protéique
Stimulation de la synthèse protéique musculaire
L’insuline est un puissant stimulateur de l’anabolisme protéique, particulièrement dans le muscle squelettique. Elle favorise la synthèse des protéines par plusieurs mécanismes. Tout d’abord, l’insuline augmente le transport des acides aminés vers l’intérieur des cellules musculaires, fournissant ainsi les éléments de base nécessaires à la synthèse protéique.
Au niveau moléculaire, l’insuline active la voie de signalisation mTOR (mammalian target of rapamycin), un régulateur clé de la synthèse protéique. L’activation de mTOR conduit à la phosphorylation de facteurs d’initiation de la traduction et de protéines ribosomales, augmentant ainsi l’efficacité de la machinerie de synthèse protéique. Cette action anabolique de l’insuline est particulièrement importante après un repas riche en protéines et en glucides.
Inhibition du catabolisme protéique
En plus de stimuler la synthèse des protéines, l’insuline inhibe leur dégradation. Elle supprime l’activité des systèmes protéolytiques cellulaires, notamment le système ubiquitine-protéasome. Cette action anti-catabolique est cruciale pour maintenir la masse musculaire, en particulier pendant les périodes de jeûne ou de stress métabolique.
L’effet net de l’insuline sur le métabolisme protéique est donc un bilan positif, favorisant l’accumulation de protéines dans les tissus. Cette action est essentielle pour la croissance, la réparation tissulaire et le maintien de la fonction musculaire. Dans des conditions pathologiques comme le diabète mal contrôlé, où l’action de l’insuline est déficiente, on observe souvent une perte de masse musculaire due à un catabolisme protéique excessif.
Dysfonctionnements de l’action de l’insuline
Résistance à l’insuline dans le diabète de type 2
La résistance à l’insuline est une caractéristique centrale du diabète de type 2. Dans cet état, les tissus cibles de l’insuline (muscles, foie, tissu adipeux) répondent de manière inadéquate à des niveaux normaux d’insuline circulante. Cette résistance se manifeste par une diminution de l’absorption du glucose par les muscles, une production hépatique de glucose excessive et une lipolyse accrue dans le tissu adipeux.
Au niveau moléculaire, la résistance à l’insuline implique des défauts dans la cascade de signalisation de l’insuline. Ces altérations peuvent survenir à différents niveaux, depuis le récepteur de l’insuline jusqu’aux effecteurs intracellulaires comme l’Akt. Des facteurs tels que l’obésité, l’inflammation chronique et le stress oxydatif contribuent au développement de la résistance à l’insuline.
Déficit en insuline dans le diabète de type 1
Le diabète de type 1 se caractérise par une destruction auto-immune des cellules bêta pancréatiques, entraînant un déficit sévère en insuline. Sans insuline endogène, les patients atteints de diabète de type 1 sont incapables de réguler efficacement leur glycémie et dépendent d’injections d’insuline exogène pour survivre.
L’absence d’insuline dans le diabète de type 1 a des conséquences métaboliques profondes. Sans le signal insulinique, les cellules ne peuvent pas absorber efficacement le glucose sanguin, conduisant à une hyperglycémie chronique. De plus, le manque d’insuline entraîne une augmentation de la lipolyse et de la cétogenèse, pouvant mener à une acidocétose diabétique potentiellement mortelle.
Syndrome métabolique et hyperinsulinémie
Le syndrome métabolique est un ensemble de conditions incluant l’obésité abdominale, l’hypertension, la dyslipidémie et l’intolérance au glucose. L’hyperinsulinémie, caractérisée par des niveaux élevés d’insuline circulante, est souvent associée à ce syndrome. Elle reflète généralement une tentative de l’organisme de compenser la résistance à l’insuline.
L’hyperinsulinémie chronique peut avoir des effets délétères, notamment en favorisant la rétention de sodium, l
‘hyperinsulinémie chronique peut avoir des effets délétères, notamment en favorisant la rétention de sodium, l’hypertension artérielle et la prolifération cellulaire anormale. De plus, elle peut exacerber la résistance à l’insuline, créant ainsi un cercle vicieux qui aggrave le syndrome métabolique.
Avancées thérapeutiques ciblant l’action de l’insuline
Analogues de l’insuline à action prolongée comme l’insuline glargine
Les analogues de l’insuline à action prolongée représentent une avancée majeure dans le traitement du diabète. L’insuline glargine, par exemple, est conçue pour avoir un profil d’action plus stable et plus long que l’insuline humaine. Sa structure moléculaire modifiée lui permet de former des micro-précipités sous-cutanés, d’où elle est libérée lentement et régulièrement.
Cette libération prolongée offre plusieurs avantages : elle réduit le risque d’hypoglycémie nocturne, permet une meilleure stabilité glycémique sur 24 heures et simplifie le schéma d’injection pour les patients. D’autres analogues à action prolongée, comme l’insuline détémir et l’insuline dégludec, ont également été développés, chacun avec ses propres caractéristiques pharmacocinétiques.
Sensibilisateurs à l’insuline comme la metformine
La metformine est le chef de file des sensibilisateurs à l’insuline et reste le traitement de première ligne pour le diabète de type 2. Son mécanisme d’action principal consiste à réduire la production hépatique de glucose, mais elle améliore également la sensibilité à l’insuline dans les tissus périphériques. Comment fonctionne exactement la metformine au niveau moléculaire ? Elle active l’AMP-kinase, une enzyme clé dans la régulation du métabolisme énergétique cellulaire.
D’autres sensibilisateurs à l’insuline, comme les thiazolidinediones, agissent en stimulant les récepteurs PPAR-γ, ce qui augmente la sensibilité à l’insuline principalement dans le tissu adipeux et les muscles. Ces médicaments peuvent être particulièrement bénéfiques chez les patients présentant une résistance à l’insuline marquée.
Thérapies basées sur les incrétines GLP-1 et DPP-4
Les thérapies basées sur les incrétines représentent une approche novatrice dans le traitement du diabète de type 2. Le GLP-1 (Glucagon-Like Peptide-1) est une hormone intestinale qui stimule la sécrétion d’insuline en réponse au glucose, inhibe la sécrétion de glucagon et ralentit la vidange gastrique. Les analogues du GLP-1, comme le liraglutide ou le sémaglutide, miment ces effets et offrent un contrôle glycémique efficace avec l’avantage supplémentaire de favoriser la perte de poids.
Les inhibiteurs de la DPP-4 (Dipeptidyl Peptidase-4), quant à eux, prolongent la demi-vie du GLP-1 endogène en empêchant sa dégradation. Ces médicaments, comme la sitagliptine ou la linagliptine, offrent une amélioration du contrôle glycémique sans risque accru d’hypoglycémie. Quelle est la différence principale entre ces deux approches ? Les analogues du GLP-1 fournissent des niveaux pharmacologiques de l’hormone, tandis que les inhibiteurs de la DPP-4 augmentent les niveaux physiologiques des incrétines endogènes.
Ces avancées thérapeutiques illustrent la complexité et la diversité des approches visant à optimiser l’action de l’insuline dans le traitement du diabète. Elles soulignent également l’importance d’une compréhension approfondie des mécanismes moléculaires régissant le métabolisme pour développer des traitements toujours plus efficaces et personnalisés.